Xiao-Bo Zhou1,
Kai-Wu Cao1,
Ling-Kun Song2,
Shuang-Qing Kou3,
Shen-Cheng Qu1,
Cong Wang1,
Ying Yu1,
Yu Liu1,
Peng-Ying Li1,
Run-Peng Lu3 ![]() ,
,
For correspondence:- Run-Peng Lu Email: lurpdlhsp@163.com
Received: 9 September 2016 Accepted: 14 December 2016 Published: 31 January 2017
Citation: Zhou X, Cao K, Song L, Kou S, Qu S, Wang C, et al. Effect of acetone extract of Rumex japonicas Houtt on hydrogen peroxide-induced apoptosis in rat myocardial cells. Trop J Pharm Res 2017; 16(1):135-140 doi: 10.4314/tjpr.v16i1.17
© 2017 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the protective effect of the acetone extract of Rumex japonicas Houtt. (AER) on rat myocardial cells.
Methods: R. japonicas was extracted with 75 % aqueous ethanol by reflux to afford total extract (TER). TER was suspended in water and then extracted with acetone to afford acetone fraction of R. japonicas (AER). High performance liquid chromatography (HPLC) combined with standard substances was carried out to analyze the major constituents of AER. Apoptosis in myocardial H9c2 cell line was induced by H2O2 (100 μmol/L). The cells were treated with AER (50, 100 and 200 μg/mL, and cell viability was evaluated by the 3-A288;4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide (MTT) assay, while oxidative stress level in H9c2 cells was evaluated by determining levels of lactate dehydrogenase (LDH), malondialdehyde (MDA), creatinine kinase (CK), superoxide dismutase (SOD), and catalase (CAT). Furthermore, apoptotic proteins (caspase-3, Bax and Bcl-2) in H9c2 cells were analyzed by using western blot assay.
Results: Results revealed that the main components of AER are aloe-emodin, rhein, emodin, chrysophanol and physcion. AER (50, 100 and 200 μg/mL) inhibited the cell viability reduction of the H9c2 cells induced by H2O2 (p < 0.05, p < 0.01, p < 0.01, respectively). AER (50, 100 and 200 μg/mL) decreased LDH and CK contents of H9c2 cells (p < 0.01). The levels of SOD (p<0.01) and CAT (p < 0.01) were increased by AER treatments (100 and 200 μg/mL); in addition, AER (50, 100 and 200 μg/mL) decreased MDA levels (p < 0.01). Besides, the present results also revealed that AER could down-regulate caspase-3 and Bax, but up-regulated Bcl-2.
Conclusion: AER alleviates apoptosis induced by H2O2 in myocardial H9c2 cells via inhibition of oxidative stress and mitochondria-mediated apoptosis. This finding suggests that AER can potentially be developed for the treatment of myocardial apoptosis.
Introduction
Plants in the Rumex genus belong to the family of Polygonaceae, and are annual or perennial herbs distributed throughout the world [1,2]. In Chinese folk medicine, plants in this genus have been used for treating various diseases from ancient times, and have medicinal functions of promoting blood circulation, removing blood stasis, detoxication, etc [2,3]. In addition, previous investigations have indicated that plants in Rumex genus contain large quantities of anthraquinone derivatives [2,4,5].
Rumex japonicus Houtt is widely distributed throughout China [6,7]. In China, the whole herb of R. japonicus has been traditionally used as an important traditional medicine for treating inflammatory diseases, tumors, infections, and constipation, etc. It is reported that R. japonicus contains a large number of anthraquinones, flavones and oxanthrones, etc [8,9]. Previous works revealed that natural extracts or monomeric compounds derived from the family of Polygonaceae are beneficial for protection of cardiac functions in many heart diseases [2,10,11]. In Chinese folk medicine, the whole herb of R. japonicas is commonly used as an ingredient of prescriptions for treating heart diseases based on the function of promoting blood circulation for removing blood stasis [2,12]. To the best of our knowledge, no experimental work on the myocardial protective effect of R. japonicas has been carried out. Thus, this study is designed to investigate the protective effects of the acetone extracts of R. japonicas (AER) on myocardial cells using a H2O2-induced myocardial apoptosis H9c2 cell model.
Methods
Cell culture
Rat ventricular cardiomyoblast H9c2 cell line was obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). The cells were cultured in the Dulbecco’s modified Eagle’s medium (DMEM) containing 10 % fetal bovine serum (FBS) at 37 °C in an incubator (Sanyo, Japan) in 5 % CO2/95 % air.
Chemicals
The DMEM media and FBS were purchased from Gibco Co. (Grand Island, NY, USA). Methyl-thiazdyldiphenyl-tetrazolium bromide (MTT), methanol (MeOH) (chromatographically pure, CP), H2O2 and dimethyl sulfoxide (DMSO) were purchased from the Sigma Ltd. Co. (St. Louis, MO, USA). Assay kits for Lactate dehydrogenase (LDH), malondialdehyde (MDA), superoxide dismutase (SOD) assay kit, creatinine kinase (CK) and catalase (CAT) were purchased from the Nanjing Jiancheng Biotech. Co. (Nanjing, China). IP cell lysis buffer, BCA protein assay kit, and Caspase-3, Bax and Bcl-2 primary antibodies were purchased from the Beyotime Co. (Haimen, China). Standard references including aloe-emodin, rhein, emodin, chrysophanol and physcion was obtained from the National Institute for the Control of Pharmaceutical and Biological Products (Beijing, China).
Plant material and AER preparation
AER was prepared according to the method described previously with minor modifications [13]. The dried whole herb of R. japonicas (purchased from the Tongrentang traditional Chinese medicine drug store, Tianjing, China) was powdered and extracted with 75 % aqueous ethanol by reflux (3 times, each for 2 h). The filtrates were combined and then evaporated at 50 °C in vacuo to afford total extract of R. japonicas (TER). Subsequently, TER was suspended in water and extracted with acetone using a 10L separating funnel, and the acetone extract evaporated at 50 °C in vacuo using a 10 L rotary evaporator (RE-5210, Shyarong Co., Shanghai, China) to afford the acetone extract of R. japonicas (AER).
HPLC analysis of AER
Methanol (MeOH) was used to dissolve the AER to obtain appropriate concentration for high performance liquid chromatography (HPLC) assays. Chemical composition analysis of AER was carried out with an Agilent Technologies 1200 system (Palo Alto, CA, USA), and performed with a Venusil XBP-C18 column (4.6 mm × 250 mm, 5 μm). The mobile phase for HPLC assay comprised of of MeOH-0.1 % phosphoric acid aqueous solution (80:20). The injection volume of sample was 10 μL, and flow rate was set at 1 mL/min. The column temperature was set at 25°C, and the detection wavelength was 254 nm.
MTT assay
H9c2 cells (5 × 104/200 μL) were cultured in 96-well plates, and then cells were treated with AER (50, 100 and 200 μg/mL) with the presence of H2O2 (100 μmo1/L) in the following day [14]. After 24 h treatment, MTT assay was carried out according to the previous report [15] and absorbance (A) was detected at 570 nm using a 96-well plate reader (BioTek Instruments, Inc, Burlington, VT, USA). Cell viability (C) was determined as in Eq 1.
C (%) = (At/Ac) 100 ……………… (1)
where At and Ac are the absorbance of treated and control samples, respectively. Each experiment was carried out in triplicate.
Determination of LDH, CK, SOD, CAT and MDA in H9c2 cells
After 24 h treatment, the H9c2 cells’ supernatants were collected to determine LDH and CK contents using commercial kits according to manufacturers’ instructions. In addition, the total cell proteins of H9c2 cells were extracted with IP cell lysis buffer, and protein concentration was analyzed using a BCA protein assay kit. Subsequently, contents of SOD, CAT and MDA were also measured using commercial kits following manufacturers’ instructions.
Western blot analysis
H9c2 cells were treated with AER (200 μg/mL) for 24 h. Then, total proteins were extracted with IP cell lysis buffer, and protein concentration was determined using the BCA protein assay kit. Subsequently, a total of 35 μg proteins were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE, 12 %), and then transferred to PVDF membranes (Boston, MA, USA). PVDF membranes were blocked with 5 % fat-free milk, and subsequently incubated with primary antibodies of caspase-3, Bax and Bcl-2, respectively. The protein bands were detected using HRP-conjugated secondary antibodies, and were visualized by the enhanced chemiluminescence reagent (Thermo Fisher Scientific, Shanghai, China). Thereafter, protein bands were quantitated using a Bio-Rad Chemi Doc XRS gel imaging system (Hercules, CA, USA).
Statistical analysis
Data are expressed as mean ± standard deviation (SD, n = 3). Statistical analysis was performed by one-way ANOVA using SPSS software (SPSS for Windows 15.0, SPSS Inc, USA). P < 0.05 was considered statistically significant.
Results
HPLC chromatograms
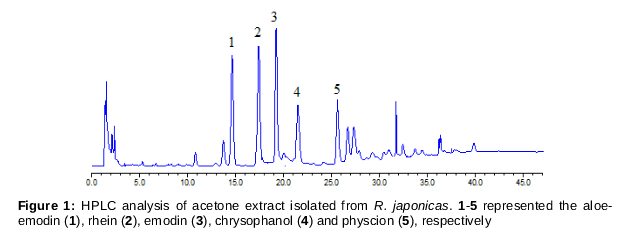
As shown in , over 10 peaks were detected within 45 min in the chromatogram map, among which 5 main compounds were identified by comparison with standard references. These 5 compounds were all anthraquinones and identified as aloe-emodin (1), rhein (2), emodin (3), chrysophanol (4) and physcion (5), respectively.
Cell viability
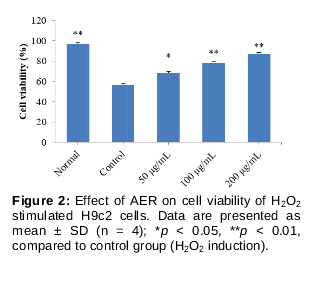
The present results () showed that after H2O2 induction (100 μmo1/L), cell viability of H9c2 cells (Control group) reduced sharply (p < 0.01), compared to normal H9c2 cells. Interestingly, AER treatments (50, 100 and 200 μg/mL) significantly inhibited the H2O2 induced cell viability reduction in H9c2 cells (p < 0.05, p < 0.01, p < 0.01, respectively), compared to control group. These results indicated that AER could enhance the cell viability of the H2O2 stimulated myocardial cell.
LDH and CK contents
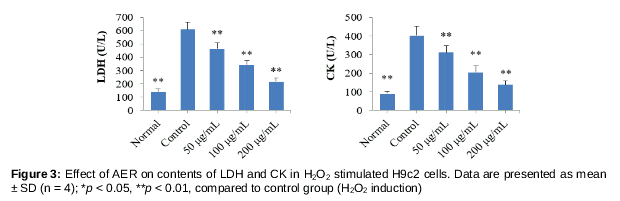
As shown in , following H2O2 stimulation, the contents of LDH and CK in H9c2 cells significantly increased (p < 0.01), compared to normal cells. After treatment with AER at concentrations of 50, 100 and 200 μg/mL, both the contents of LDH and CK significantly decreased (p < 0.01), compared to control group. These results indicate that AER alleviated leakages of CK and LDH from cytoplasm due to H2O2-induced cell membrane damage.
SOD, CAT and MDA levels
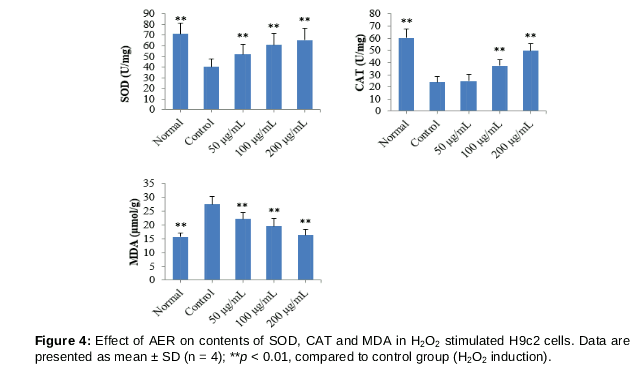
Levels of SOD, CAT and MDA in H9c2 cells were represented in . After stimulation with H2O2, SOD and CAT levels in H9c2 cells sharply decreased (p < 0.01), whereas the MDA content significantly increased (p < 0.01), compared to normal H9c2 cells. Interestingly, the levels of SOD and CAT in H9c2 cells were significantly increased by treating AER (100 and 200 μg/mL, p < 0.01), compared to control group. In addition, the levels of MDA in AER (50, 100 and 200 μg/mL) treated group decreased significantly (p < 0.01, p < 0.01, p < 0.01), compared to control group.
Apoptosis
Apoptotic proteins’ expressions (including caspase-3, Bcl-2 and Bax) were determined by western blot assays. Compared to normal H9c2 cells, after treatment with H2O2, expressions of caspase-3 and Bax up-regulated, whereas the Bcl-2 expression down-regulated. However, expressions of caspase-3 and Bax down-regulated after treatment with AER (200 μg/mL), whereas the Bcl-2 expression up-regulated, compared to control group.
Discussion
Currently, cardiac diseases have become one of the leading causes of death worldwide, and the prevalence of cardiac diseases are growing rapidly [16]. In addition, ischemic coronary heart diseases, such as myocardial infarction, could result in myocardial apoptosis, leading to cardiac failure [16,17]. Therefore, inhibiting myocardial apoptosis is beneficial for treating ischemic coronary heart diseases.
The myocardial cell function is dependent on O2 consumption by mitochondria and electron transport chain [18,19]. Previous reports indicated that myocardial ischemia is mainly caused by imbalance of demand and supply of oxygen to heart [14,20]. Furthermore, increasing researches have demonstrated that oxidative stress is the crucial cause of pathogenesis of myocardial apoptosis [21]. Oxidative stress could result in over-produced oxygen free radical (ROS) which could damage cell membrane, enzymes, and electron transport chain, inducing cell apoptosis. According to previous investigations, H2O2 induced myocardial apoptosis H9c2 cell model is commonly used to investigate pathological process and molecular mechanism of myocardial ischemia injury [20,22]. The results demonstrate that AER can alleviate the myocardial apoptosis induced by H2O2; furthermore, AER can also alleviate the leakages of CK and LDH from the cytoplasm due to H2O2 induced cell membrane damage. In oxidative stress, MDA is a major end-product of lipid peroxidation, and is considered as an indicator of tissue damage [23]. In contrast, GAT and SOD, the crucial anti-oxidative enzymes, are considered as antioxidant defense system in body which plays crucial roles in suppressing oxidative stress [24]. This work indicates that AER increase the levels of GAT and SOD, but decreases MDA level in H2O2 induced H9c2 cell. These results suggest that AER inhibits oxidative stress level in H2O2-induced H9c2 cells.
It is reported that inhibition of myocardial apoptosis may be beneficial for treatment and prognosis of various ischemic heart diseases. Caspase family proteins play key roles in regulating apoptosis induced by different stimulus such as oxidative stress, and caspase-3 is the most important executor in the apoptotic process [25]. Furthermore, Bax is a pro-apoptotic protein in the mitochondria-mediated apoptosis, whereas Bcl-2 is an anti-apoptotic protein [24]. In the present study, AER down-regulated the caspase-3 and Bax, but up-regulated Bcl-2, indicating that AER suppresses apoptosis in cardiomyocytes.
Conclusion
The findings of the present investigation demonstrate that the acetone extract of R. japonicas (AER) alleviates myocardial apoptosis in H2O2-induced H9c2 cell. The probable mechanism might be related to the inhibition of oxidative stress and mitochondria-mediated apoptosis. These findings suggest that AER can be further developed for the treatment of myocardial disease.
Declarations
Acknowledgement
References
Archives


News Updates